"Большая Советская Энциклопедия (МХ)" - читать интересную книгу автора (БСЭ БСЭ)
 |
Большая Советская Энциклопедия (МХ)
Мхи
Мхи, наземные или реже пресноводные автотрофные растения, объединяемые в отдел
Антоцеротовые М. (Anthocerotae) — своеобразная группа из 2 семейств с 6 родами, объединяющими свыше 300 видов, распространённых преимущественно в тропиках, в СССР 4—5 видов. Слоевище у них обычно лопастное или розетковидное с погружёнными в него антеридиями и архегониями. Спорогон, как правило, вытянутый, стручковидный, способный удлиняться благодаря
Печёночные М., или печёночники (Hepaticae), подразделяются на 2 подкласса: маршанциевые и юнгерманниевые. В отличие от других М., у печёночников протонема слабо развита, недолговечна и на ней обычно развивается одно слоевищное или листостебельное растение дорзо-вентральной структуры; в клетках гаметофита содержатся одно или несколько масляных телец постоянной формы; ризоиды одноклеточные. Спорогон чаще без хлоропластов, лишён колонки, и на его стенках отсутствуют устьица. У некоторых маршанциевых обнаруживается сходство с антоцеротовыми, поэтому антоцеротовых рассматривали как подкласс печёночных М. Гаметофит печёночников отличается большим разнообразием, спорогоны однотипны. Маршанциевые печёночники (Marchantiidae) — слоевищные формы, обычно в виде дихотомически ветвящихся маленьких розеток или крупных лентовидно стелящихся слоевищ, прикрепляющихся к почве ризоидами; мужские и женские гаметангии погружены в ткань слоевища или приподняты на особых подставках. Преимущественно напочвенные растения, редко скальные или водные, распространены по всему земному шару, особенно богато представлены в тропиках. Известно 16 семейств с 35 родами, объединяющими около 420 видов, включая ископаемую Naiadita из мезозоя. Наиболее примитивны М. из порядка сферокарповых с просто устроенным слоевищем, иногда с масляными тельцами в особых клетках, с однослойной стенкой спорогона и с питательными клетками вместо элатер. Для М. порядка маршанциевых характерно сложное строение слоевища с разделением на основную и ассимиляционную ткань, с масляными тельцами в особых клетках и со склеренхимными волокнами (у некоторых видов). Слоевища часто с воздушными камерами и воздушными устьицами, с нижней стороны с брюшными чешуйками и одноклеточными гладкими и язычковыми ризоидами. Спорогон обычно более сложного строения. Среди них риччиевые рассматриваются как вторично упрощённые, с гаметангиями и спорогоном, развивающимся внутри мелких, чаще розетковидных слоевищ. Юнгерманниевые печёночники (Jungermanniidae) включают группу слоевищных и олиственных растений, в вегетативных клетках которых, как правило, по нескольку масляных телец; ризоиды гладкие, одноклеточные. Эпифитные, напочвенные и наскальные растения особенно богато представлены в субтропиках и тропиках. 45 семейств объединяют свыше 240 родов, включающих около 9 тыс. видов. У представителей порядка метцгериевых слоевище одно- или многослойное, недифференцированное (у некоторых с брюшными или спинными, иногда листовидными чешуйками и стеблями с листовидными лопастями); коробочка двух- или многослойная. Растения порядка гапломитриевых прямостоячие, с трёхрядным листорасположением и коробочками, имеющими однослойные боковые стенки. Наиболее многочисленны и разнообразны М. порядка юнгерманниевых, для которых характерны 2 ряда цельных или лопастных боковых листьев без жилки и более мелкие (иногда отсутствуют) брюшные листья — так называемые амфигастрии; стенки коробочки многослойные.
Лиственные М., или листостебельные М. (Musci), подразделяются на 3 подкласса: сфагновые, андреевые и бриевые. Растения радиально-, редко двусторонне-симметричные, спиральноолиственные; листья цельные, зубчатые или по краю реснитчатые, однослойные, реже дву- или многослойные, с жилкой или без жилки; с многоклеточными, ветвистыми ризоидами. Спорогоны верхушечные или боковые, разнообразные по форме, строению и окраске. Около 15 тыс. видов (по др. данным, до 25 тыс. видов), объединяемых в 660 родов. Сфагновые, или торфяные, М. имеют крупные, прямостоячие стебли с пучковидно расположенными ветвями, на верхушке собранными в головку; беловато-зелёные, жёлтые, бурые или красноватые. Листья однослойные, веточные и стеблевые, без жилки, из чередующихся хлорофиллоносных и водоносных клеток. Спорогон состоит из шаровидной коробочки с крышечкой и недоразвитыми устьицами, без ножки, со стопой, вросшей в безлистную удлинённую архегониальную веточку стебля. Споровый мешок сводообразно покрывает верхнюю часть колонки. Распространены по всему земному шару: преобладают на болотах, в тундре, во влажных лесах. Андреевые М. — мелкие красно-бурые (до чёрных), с однослойными листьями; растут на скалах подушечками. Коробочка удлинённо-яйцевидная, на ложноножке, раскрывается 4—8 створками, без устьиц, со споровым мешком в виде свода над верхней частью колонки. Распространены высоко в горах, а также в северных широтах. Бриевые М. — наиболее обширная, разнообразная группа. Коробочка обычно на настоящей ножке, с устьицами, чаще после опадения крышечки открывается наверху широким отверстием — устьем. У многих по краю устья образуется 1—2 ряда выростов — зубцов (называемых простым или двойным перистомом), которые служат для рассеивания спор. Колонка, как правило, пронизывает споровый мешок. (Цикл развития см.
М. распространены повсеместно. Поселяются повсюду, кроме морей, засолённых почв и местообитаний, скрытых под ледниками или подверженных сильной эрозии. Антоцеротовые и печёночные М. распространены в основном в странах с тропическим или умеренным влажным климатом; только немногие приспособились к произрастанию в сухих местах. Лиственные М. растут почти повсеместно, однако наибольшего развития достигают на увлажнённых местах, в лесах, тундре; на болотах М. образуют основную массу торфяных залежей. Интенсивно развиваясь, М. способствуют заболачиванию почв, ухудшают качество лугов и др. с.-х. угодий. В связи с антибиотическими свойствами некоторых М. их применяют иногда как перевязочный материал. Используют в качестве подстилки для скота (преимущественно сфагновые М.), в строительной технике для изготовления плит и др.
 |
Атоцеротовые мхи: 1 — Anthoceros punctatus. Печёночные мхи: 2 — Riccia glauca, 3 — Riccia fluitans.
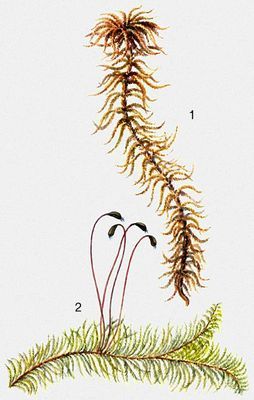 |
Листостебельные мхи: 1 — Sphagnum fuxum, 2 — Ptilium crista-castrensis.
 |
Печёночные мхи: 1 — Plagiochila asplenioides. Листостебельные мхи: 2 — Sphagnum magellanicum, 3 — Sphagnum palustre.
 |
Печёночные мхи: 1 — Marchantia polymorpha. Листостебельные мхи: 2 — Dicranum scoparium, 3 — Sphagnum recurvum, 4 — Splachnum rubrum, 5 — Polytrichum commune, 6 — Fontinalis antipyretica, 7 — Bryum marratii, 8 — Rhytidiadelphus triquetrus.
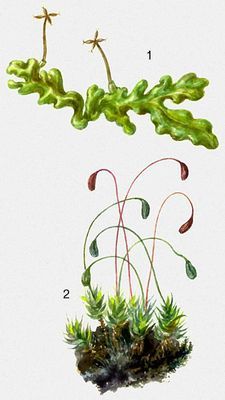 |
Печёночные мхи: 1 — Riccardia pinguis. Листостебельные мхи: 2 — Funaria hygrometrica.
 |
Печёночные мхи: Conocephalum conicum.
| © 2024 Библиотека RealLib.org (support [a t] reallib.org) |