"Большая Советская Энциклопедия (ОК)" - читать интересную книгу автора (БСЭ БСЭ)
Окисление биологическое
Окисле'ние биологи'ческое, совокупность реакций окисления, протекающих во всех живых клетках. Основная функция О. б. — обеспечение организма энергией в доступной для использования форме. Реакции О. б. в клетках катализируют ферменты, объединяемые в класс оксидоредуктаз. Изучение окисления в организме было начато в 18 в. А. Лавуазье; в дальнейшем значительный вклад в исследование О. б. (его локализация в живых клетках, связь с др. процессами обмена веществ, механизмы ферментативных окислительно-восстановительных реакций, аккумуляция и превращение энергии и др.) внесли О. Варбург, Г. Виланд (Германия), Д. Кейлин, Х. Кребс, П. Митчелл (Великобритания), Д. Грин, А. Ленинджер, Б. Чанс, Э. Рэкер (США), а в СССР — А. Н. Бах,В. И. Палладин, В. А. Энгельгардт, С. Е. Северин,В. А. Белицер, В. П. Скулачев и др.
О. б. в клетках связано с передачей т. н. восстанавливающих эквивалентов (ВЭ) — атомов водорода или электронов — от одного соединения — донора, к другому — акцептору. У аэробов— большинства животных, растений и многих микроорганизмов — конечным акцептором ВЭ служит кислород. Поставщиками ВЭ могут быть как органические, так и неорганические вещества (см. таблицу).
Классификация организмов по источнику энергии и восстанавливающих эквивалентов
Тип организмов
Источник энергии
Окисляемое соединение (поставщик восстанавливающих эквивалентов)
Неорганические соединения (Н2О, H2S, S) Органические соединения Неорганические соединения (H2, S, H2S, NH3, Fe2+) Органические соединения
Зелёные клетки высших растений, синезелёные водоросли, фотосинтезирующие бактерии Несерные пурпурные бактерии Водородные, серные, денитрифицирующие бактерии, железобактерии Животные, большинство микроорганизмов, нефотосинтезирующие клетки растений
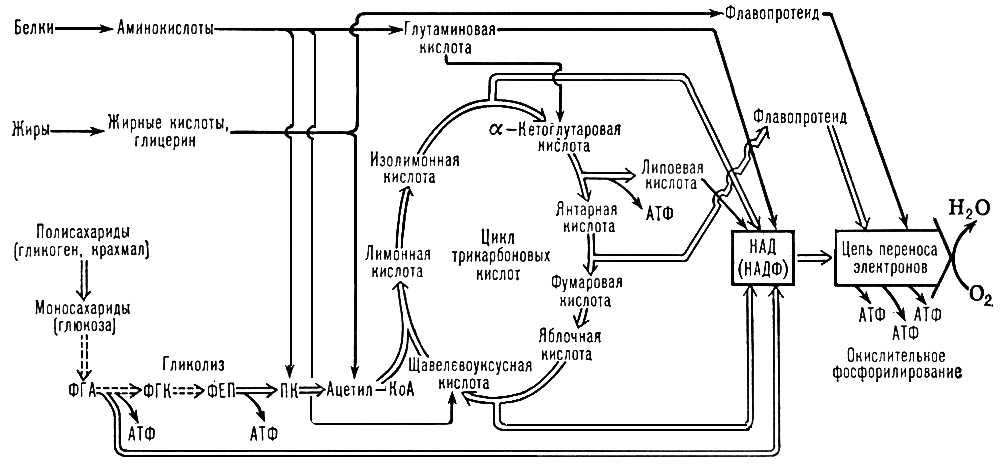
Основной путь использования энергии, освобождающейся при О. б., — накопление её в молекулах аденозинтрифосфорной кислоты (АТФ) и др. макроэргических соединений.О. б., сопровождающееся синтезом АТФ из аденозиндифосфорной кислоты (АДФ) и неорганического фосфата, происходит при гликолизе, окислении a-кетоглутаровой кислоты и при переносе ВЭ в цепи окислительных (дыхательных) ферментов, обычно называют окислительным фосфорилированием (см. схему).
В процессе дыхания углеводы, жиры и белки подвергаются многоступенчатому окислению, которое приводит к восстановлению основных поставщиков ВЭ для дыхательных флавинов, никотинамидадениндинуклеотида (НАД), никотинамидадениндинуклеотидфосфата (НАДФ) и липоевой кислоты. Восстановление этих соединений в значительной мере осуществляется в трикарбоновых кислот цикле, которым завершаются основные пути окислительного расщепления углеводов (оно начинается с гликолиза), жиров и аминокислот. Помимо цикла трикарбоновых кислот, некоторое количество восстановленных коферментов— ФАД (флавинадениндинуклеотида) и НАД — образуется при окислении жирных кислот, а также при окислительном дезаминировании глутаминовой кислоты (НАД) и в пентозофосфатном цикле (восстановленный НАДФ).
Соотношение и локализация различных механизмов О. б. В расчёте на 1 молекулу глюкозы гликолиз даёт 2 молекулы АТФ, а фосфорилирование в дыхательной цепи — 34 молекулы АТФ. Гликолиз, цикл трикарбоновых кислот и дыхательная цепь функционируют, по-видимому, в клетках всех эукариотов. Окисление жирных кислот у позвоночных поставляет половину энергии, потребляемой печенью, почками, мышцей сердца и покоящимися скелетными мышцами; в клетках мозга оно практически не происходит. Окисление по пентозофосфатному пути активно в печени и лактирующих молочных железах, но незначительно в сердечной и скелетных мышцах.
В жидкой фазе цитоплазмы растворены все ферменты гликолиза. Внутренние мембраны митохондрий, мембраны хлоропластов (тилакоидов) и клеточные мембраны бактерий содержат фосфорилирующие цепи переноса электронов. В матриксе митохондрий локализовано окисление жирных кислот, ферменты цикла трикарбоновых кислот и глутаматдегидрогеназа. Во внутренней мембране митохондрий находятся ферменты, окисляющие янтарную и b-оксимасляную кислоты, во внешней — ферменты, участвующие в обмене аминокислот: моноаминоксидаза и кинуренингидроксилаза. В особых органоидах клетки, т. н. пероксисомах, или микротельцах, вклад которых в суммарное поглощение О2 может достигать в печени 20%, находится флавиновая оксидаза, окисляющая аминокислоты, гликолевую кислоту и др. субстраты с образованием перекиси водорода, которая затем разлагается каталазой или используется пероксидазами в реакциях окисления. В мембранах эндоплазматической сети клетки локализованы гидроксилазы и оксигеназы, организованные в короткие нефосфорилирующие цепи переноса электронов.
Окислительные реакции не всегда сопровождаются накоплением энергии; в ряде случаев они несут функции превращения веществ (например, окисление при образовании жёлчных кислот, стероидных гормонов, на путях превращения аминокислот и др.). При окислении происходит обезвреживание чужеродных и ядовитых для организма веществ (ароматических соединений, недоокисленных продуктов дыхания и др.). О. б., не сопряжённое с накоплением энергии, называется свободным окислением. Его энергетический эффект — образование тепла. По-видимому, система переноса электронов, осуществляющая окислительное фосфорилирование, способна переключаться на свободное окисление при увеличении потребности организма в тепле (у гомойотермных животных).
Механизм использования энергии окисления. Долгое время оставался неясным вопрос о механизме преобразования энергии, освобождающейся при переносе ВЭ по цепи окислительных ферментов. Согласно т. н. хемиосмотической теории, развитой в 60-х гг. 20 в. (английский биохимик П. Митчелл и др.), энергия сначала используется для создания электрического поля («+» с одной стороны мембраны и «–» с другой) и разности концентраций ионов Н+ по разные стороны мембраны. Оба фактора (электрическое поле и разность концентраций) могут служить движущей силой для действия фермента АТФ-синтетазы, осуществляющей синтез АТФ. Часть энергии поля может быть прямо использована клеткой для переноса ионов через мембрану, восстановление переносчиков электронов, образования тепла без промежуточного участия АТФ.
Эволюция энергообеспечения в живой природе. Древнейшие организмы, как полагают, существовали в первичной бескислородной атмосфере Земли и были анаэробами и гетеротрофными организмами.Обеспечение клеток энергией шло за счёт процессов типа гликолиза. Возможно, существовал механизм окисления, известный у некоторых современных микроорганизмов: ВЭ передаются через дыхательную цепь на нитрат (NO–3) или на сульфат (SO– –4). Принципиально важным этапом оказалось возникновение у древних одноклеточных организмов механизма фотосинтеза, с которым связывают появление кислорода в атмосфере Земли. В результате стало возможным использование O2, обладающего высоким окислительно-восстановительным потенциалом, в качестве конечного акцептора электронов в дыхательной цепи. Реализация этой возможности произошла при появлении специального фермента — цитохромоксидазы, восстанавливающей О2, и привела к возникновению биохимического дыхательного аппарата современного типа. Обеспечение энергией у всех аэробов (их клетки содержат митохондрии) основано на таком дыхании. Вместе с тем клетки сохранили ферментный аппарат гликолиза. Образуемая в ходе последнего пировиноградная кислота окисляется далее в цикле трикарбоновых кислот, который, в свою очередь, питает дыхательную цепь электронами. Т. о., эволюция энергетического обмена шла, по-видимому, по пути использования и надстройки уже имевшихся ранее механизмов энергообеспечения. Наличие в клетках ныне существующих организмов биохимических систем гликолиза (в цитоплазме), дыхания (в митохондриях), фотосинтеза (в хлоропластах), а также поразительное сходство механизмов превращения энергии в этих органеллах и в микроорганизмах нередко рассматривают как свидетельство возможного происхождения хлоропластов и митохондрий от древних микроорганизмов-симбионтов. См. также Аденозинфосфорные кислоты,Биоэнергетика,Брожение,Дыхание,Митохондрии,Фотосинтез и лит. при этих статьях.
Лит.: Ленингер А., Превращение энергии в клетке, в кн.: Живая клетка, пер. с англ., М., 1962; Скулачев В. П., Аккумуляция энергии в клетке, М., 1969; его же, Трансформация энергии в биомембранах, М., 1972; Малер Г. и Кордес Ю., Основы биологической химии, пер. с англ., М., 1970, гл. 15; Леви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971, гл. 12; Ясайтис А. А., Превращение энергии в митохондриях, М., 1973; Ленинджер А., Биохимия, пер. с англ., М., 1974.
С. А. Остроумов.
Пути образования АТФ при хемоорганотрофном типе энергетического обмена. ФГА — 3-фосфоглицериновый альдегид; ФГК — 3-фосфоглицериновая кислота; ФЕП — фосфоенолпировиноградная кислота; ПК — пировиноградная кислота; Ацетил-КоА — ацетил-кофермент А. Количественные соотношения отдельных путей биологического окисления показаны одинарными и двойными стрелками.